

miR-132, an experience-dependent microRNA, is essential for visual cortex plasticity
Mellios N, Sugihara H, Castro J, Banerjee A, Le C, Kumar A, Crawford B, Strathmann J, Tropea D, Levine SS, Edbauer D, Sur M
04.09.2011
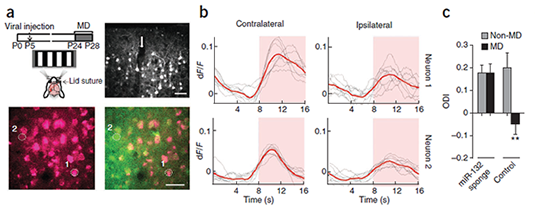
In vivo inhibition of miR-132 in V1 neurons disrupts their ocular dominance plasticity. (a) Top left, schematic of experimental design. Top right, confocal microscopy images showing mCherry expression in cortical layers 1–3 at P28 following neonatal injection (arrow) of mCherry–miR-132 sponge–expressing lentivirus. Bottom, two-photon microscopy image showing mCherry expression alone (left) and its overlay with OGB (green) fluorescence (right) in an injected mouse that underwent monocular deprivation; selected neurons used in b are circled and numbered. Scale bars represent 40 μm. (b) Example of contralateral eye– and ipsilateral eye–driven calcium responses of the neurons shown in a that express high (neuron 1) or low (neuron 2) levels of mCherry–miR-132 sponge. Pink shaded area indicates period with visual stimulus. Calculated ODIs were 0.73 (neuron 1) and 0.55 (neuron 2). (c) ODI values (ODI = (contralateral – ipsilateral) / (contralateral + ipsilateral); mean ± s.e.m.) derived from peak visual responses obtained by two-photon calcium imaging in mice injected with mCherry–miR-132 sponge–expressing lentivirus. Black bars indicate mice subjected to 4 d of monocular deprivation and light gray bars indicate mice that were not subjected to monocular deprivation (miR-132 sponge non–monocular deprivation, five mice, 290 neurons; miR-132 sponge monocular deprivation, four mice, 232 neurons; control non–monocular deprivation; three mice, 120 neurons; control monocular deprivation, three mice, 177 neurons). The ODI did not shift following monocular deprivation in mice that were injected with miR-132 sponge, but not control, virus. A significantly larger ocular dominance shift was seen in control monocularly deprived mice than in any of the other conditions (**P < 0.01, Mann-Whitney test comparing neurons; P < 0.05 treating each mouse as a single datum).
ABSTRACT
Using quantitative analyses, we identified microRNAs (miRNAs) that were abundantly expressed in visual cortex and that responded to dark rearing and/or monocular deprivation. The most substantially altered miRNA, miR-132, was rapidly upregulated after eye opening and was delayed by dark rearing. In vivo inhibition of miR-132 in mice prevented ocular dominance plasticity in identified neurons following monocular deprivation and affected the maturation of dendritic spines, demonstrating its critical role in the plasticity of visual cortex circuits.
If the role of miR-132 in plasticity extends to other cortical areas, these findings may have important functional implications for schizophrenia and Alzheimer's disease, where miR-132 downregulation has been reported previously (Kim AH et al, Schizophr. Res. 2010 and Cogswell JP et al, J. Alzheimers. Dis. 2008).

